SAIZONOU Marie-Ange: Understanding of development and differentiation of the epithelium on Urinary Collecting System (UCS) (urothelium) in human embryonic metanephros; comparison with the urinary duct and bladder
Return of the intestinal loop and superior mesenteric artery to the abdominal coelom after physiological umbilical herniation 生理的臍帯ヘルニアの還納過程における3次元的解析 掛谷真樹
56.Kakeya M, Matsubayashi J, Kanahashi T, Männer J, Yamada S, Takakuwa T. The return process of physiological umbilical herniation in human fetuses: the possible role of the vascular tree and umbilical ring. J Anatomy 2022, 241(3), 846-859. https://doi.org/10.1111/joa.13720
Fixation of the colorectum in the abdominal coelom during normal human development ヒト胎児期における大腸の還納、固定時期の解析 黄 柔婷
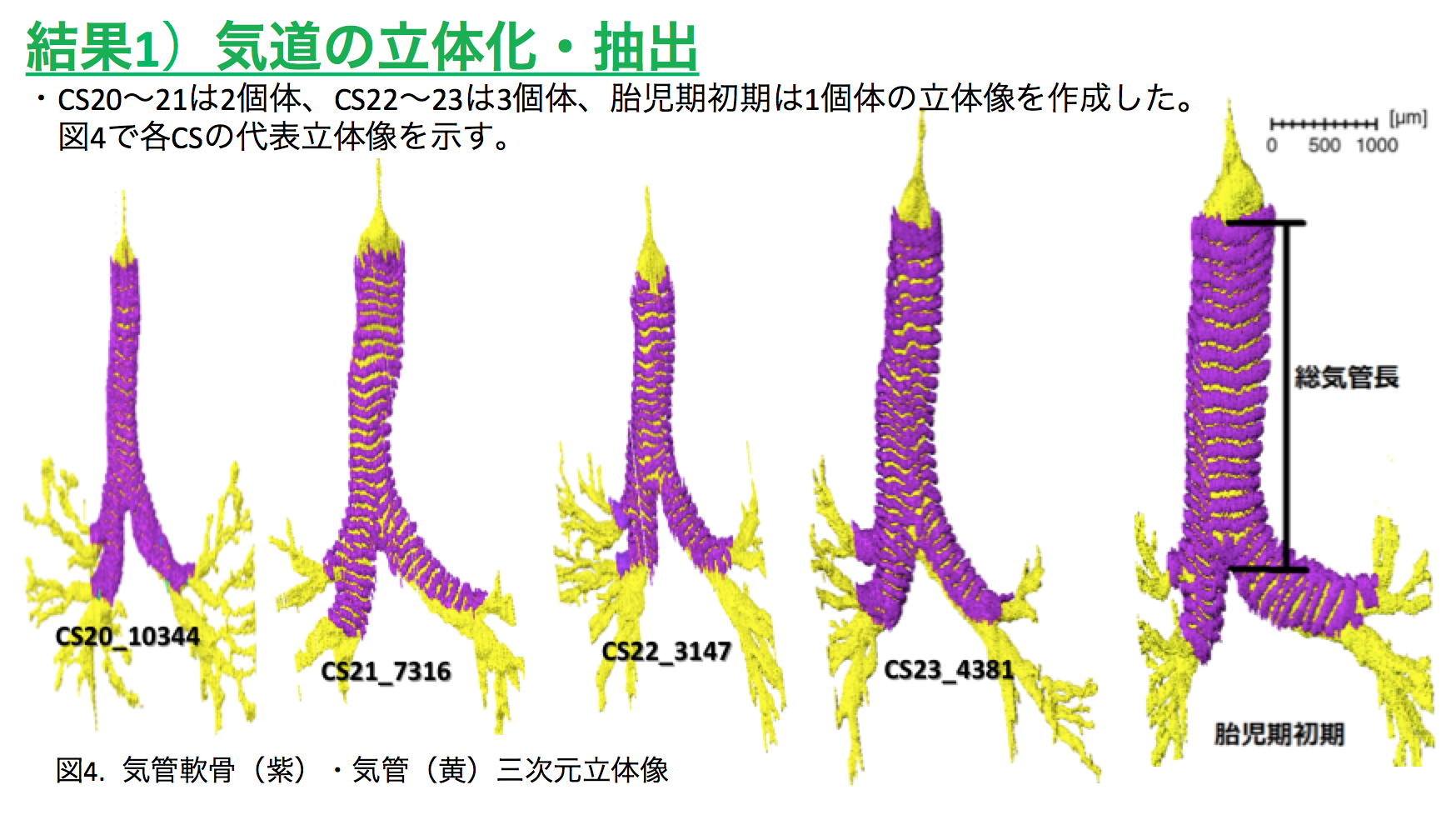
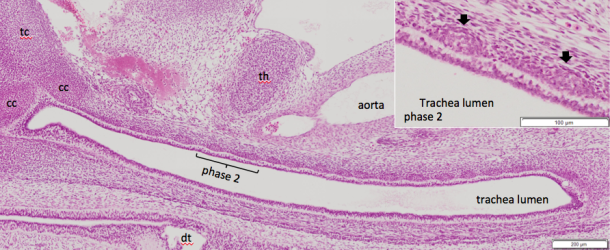
54. Yamazaki Y, Kanahashi T, Yamada S, Männer J, Takakuwa T. Three-dimensional analysis of human laryngeal and tracheobronchial cartilages during the late embryonic and early fetal period. Cells Tissues Organs, 2021 in press
Abstract
Laryngeal and tracheobronchial cartilages are present as unique U-shaped forms around the respiratory tract and contribute to the formation of rigid structures required for the airway. Certain discrepancies still exist concerning cartilage formation in humans. To visualize the accurate timeline of cartilage formation, tracheobronchial and laryngeal cartilages were 3D reconstructed based on serial tissue sections during the embryonic period (Carnegie stage [CS] 18–23) and early fetal period (crown rump length [CRL] = 35–45 mm). The developmental phases of the cartilage were estimated by histological studies, which were performed on the reconstructed tissue sections. The hyoid greater horns were recognizable at CS18 (phase 2). Fusion of 2 chondrification centers in the mid-sagittal region was observed at CS19 in the hyoid bone, at CS20 in the cricoid cartilage, and in the specimen with CRL 39 mm in the thyroid cartilage. Phase 3 differentiation was observed at the median part of the hyoid body at CS19, which was the earliest among all other laryngeal and tracheobronchial cartilages. Most of the laryngeal cartilages were in phase 3 differentiation at CS22 and in phase 4 differentiation at CS23. The U-shaped tracheobronchial cartilages with phase 2 differentiation covered the entire extrapulmonary region at CS20. Phase 3 differentiation started on the median section and propagates laterally at CS21. The tracheobronchial cartilages may form simultaneously during the embryonic period at CS22-23 and early fetal periods, similar to adults in number and distribution. The spatial propagation of the tracheal cartilage differentiation provided in the present study indicates that cartilage differentiation may have propagated differently on phase 2 and phase 3. This study demonstrates a comprehensible timeline of cartilage formation. Such detailed information of the timeline of cartilage formation would be useful to improve our understanding of the development and pathophysiology of congenital airway anomalies.
pSP-IZ の厚さは 3D で表示され、中外側領域で大きかった。CRL 64 mm)に、厚い領域は、島の原基の周りの外側、上部、および後部領域に拡大しました。
層構造は、発達初期に厚くなるのではなく、薄く成長して広がった。
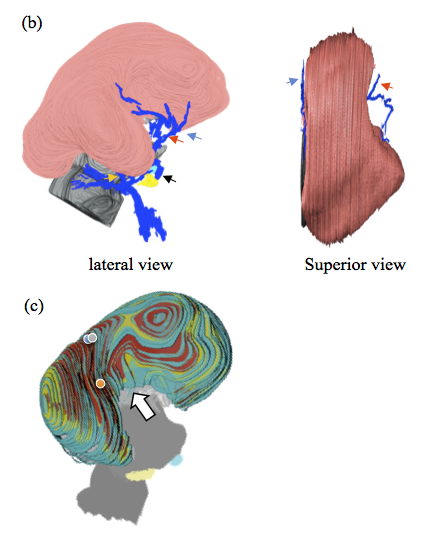
Terashima, M., Ishikawa A., Männer J., Yamada S.&Takakuwa T. (2021) Early development of the cortical layers in the human brain. Journal of Anatomy, 239, 1039–1049. https://doi.org/10.1111/joa.13488
Abstract
The cortical plate (CP) first appears at seven postconceptional weeks (pcw), when it splits the preexisting preplate into two layers, the marginal zone and the presubplate (pSP). Although three-dimensional (3D) analysis using fetal magnetic resonance imaging and two-dimensional tissue observations have been reported, there have been no studies analyzing the early development of the layer structure corresponding to the pSP stage in 3D. Here, we reconstructed 3-D models of the brain with a focus on the cortical layers in pSP stage. To achieve this, we digitized serial tissue sections of embryos between CS20 and CS23 from the Kyoto Collection (n = 7, approximately 7–8.5 pcw), and specimens at early fetal phase from the Blechschmidt Collection (n = 2, approximately 9.5–12 pcw, crown rump length [CRL] 39 and 64 mm). We observed tissue sections and 3D images and performed quantitative analysis of the thickness, surface area, and volume. Because the boundary between pSP and the intermediate zone (IZ) could not be distinguished in hematoxylin and eosin-stained sections, the two layers were analyzed together as a single layer in this study. The histology of the layers was observed from CS21 and became distinct at CS22. Subsequently, we observed the 3-D models; pSP-IZ was present in a midlateral region of the cerebral wall at CS21, and an expansion centered around this region was observed after CS22. We observed it over the entire cerebral hemisphere at early fetal phase (CRL 39 mm). The thickness of pSP-IZ was visible in 3D and was greater in the midlateral region. At the end of the pSP stage (CRL 64 mm), the thick region expanded to lateral, superior, and posterior regions around the primordium of the insula. While, the region near the basal ganglia was not included in the thickest 10% of the pSP-IZ area. Middle cerebral artery was found in the midlateral region of the cerebral wall, near the area where pSP-IZ was observed. Feature of layer structure growth was revealed by quantitative assessment as thickness, surface area, and volume. The maximum thickness value of pSP-IZ and CP increased significantly according to CRL, whereas the median value increased slightly. The layer structure appeared to grow and spread thin, rather than thickening during early development, which is characteristic during pSP stages. The surface area of the cerebral total tissue, CP, and pSP-IZ increased in proportion to the square of CRL. The surface area of CP and pSP-IZ approached that of the total tissue at the end of the pSP stage. Volume of each layer increased in proportion to the cube of CRL. pSP-IZ and CP constituted over 50% of the total tissue in volume at the end of the pSP stages. We could visualize the growth of pSP-IZ in 3D and quantify it during pSP stage. Our approach allowed us to observe the process of rapid expansion of pSP-IZ from the midlateral regions of the cerebral wall, which subsequently becomes the insula.







 共同研究者の勝部先生の下記論文が2021年度の日本形成外科学会学術奨励賞基礎部門に選ばれました。おめでとうございます。
共同研究者の勝部先生の下記論文が2021年度の日本形成外科学会学術奨励賞基礎部門に選ばれました。おめでとうございます。